

Per competizione interspecifica s'intende la riduzione della fecondità, della sopravvivenza oppure dell'accrescimento d'una popolazione a causa della presenza d'altre specie interferenti.
Rispetto a come si verifica la competizione intraspecifica le modalità sono le stesse, ma si tratta comunque di individui appartenenti a specie differenti e che, pertanto, presentano caratteristiche riproduttive e di sfruttamento delle risorse tra loro diverse. Ciò fa sì che le conseguenze della competizione intraspecifica siano differenti da quella interspecifica. In effetti, gli effetti provocati dalla competizione interspecifica possono influenzare le dinamiche delle popolazioni coinvolte dalla competizione il che, alla fine, può determinare conseguenze anche a livello evolutivo.
Gli esperimenti di Gause
Nel libro di Georgyi Frantsevitch Gause del 1934 vennero illustrati alcuni classici esperimenti con due specie di protozoi: Paramecium aurelia e Paramecium caudatum.
Questi organismi unicellulari vennero posti in terreni di coltura costituiti da 5 cc di soluzione salina (soluzione salina di Osterhout) cui era stata aggiunta una quantità nota di Bacillus pyocyaneus come fonte di nutrimento.
Ponendo in colture separate le due specie, e rinnovando periodicamente il terreno, si ottennero curve di accrescimento approssimativamente sigmoidali con il conseguimento d'uno stato stazionario. Ponendo le due specie insieme in uno stesso terreno di coltura si vide che mentre Paramecium aurelia manteneva ancora un accrescimento logistico, la popolazione di Paramecium caudatum, dopo un certo numero di giorni (circa 8), cominciò a diminuire fino a scomparire del tutto.
Da ciò si può dedurre che una delle due specie è riuscita a competere meglio per le risorse causando l'estinzione di quella concorrente.
Il principio d'esclusione competitiva
Dall'esempio riportato e da altri che sono stati descritti, Gause formulò il principio d'esclusione competitiva (o principio di Gause). Tale principio afferma che se due specie coesistono in un medesimo ambiente ciò avviene in ragione del fatto che esse presentano nicchie ecologiche separate. Qualora, però, le due specie presentino nicchie sovrapposte, allora una delle due specie prenderà il sopravvento sull'altra fino ad eliminarla.
Tale principio, comunque, può risultare difficilmente applicativo in quanto in natura si possono trovare specie simili che coesistono. Spesso la coesistenza è garantita dalla presenza di nicchie ecologiche non completamente sovrapposte (le specie in questione possono presentare, infatti, differenze lievi a livello di dieta o di habitat). In conseguenza di ciò nasce il quesito su quanto due nicchie debbano essere separate affinché la coesistenza sia permessa.
Le prime risposte a tale quesito derivarono dall'applicazione del modello della competizione interspecifica di Lotka-Volterra, così chiamato dal nome dei due ideatori che lo svilupparono in parallelo.
La mutua esclusione
La mutua esclusione è un tipo di relazione interspecifica che ha luogo tra due specie incompatibili tali che sia completamente impossibile per queste vivere insieme. All'apparire di una specie, l'altra è assente e viceversa, per cause attribuibili al comportamento competitivo di entrambe.
In natura questo fenomeno si verifica generalmente quando una delle due specie altera l'habitat in modo tale da rendere impossibile la vita all'altra, potendo in effetti quest'ultima vivere perfettamente nello stesso habitat in assenza dell'altra specie.
Il modello di Lotka e Volterra della competizione interspecifica
Il modello di Lotka e Volterra è un modello matematico descrivente la dinamica di popolazioni che competono per le medesime risorse. La base del modello è costituita dall'equazione logistica che, in ecologia, viene espressa nella seguente forma:
- (eq. 1,0)

dove N rappresenta il numero d'individui d'una popolazione ad un tempo t, r è il tasso intrinseco d'aumento della popolazione e K rappresenta la capacità portante (ovvero la densità di popolazione presso cui i tassi di mortalità e natalità risultano essere uguali).
Nel modello di Lotka e volterra il termine tra parentesi dell'equazione logistica viene sostituito con un altro che incorpora la competizione interspecifica.
Modello a due specie
Siano due differenti popolazioni con dinamica logistica, ed .
Nell'equazione logistica il termine rappresenta la competizione intraspecifica. Se N tende a K allora il rapporto tende ad 1 ed il termine a 0, condizione di equilibrio stabile.
Per esprimere la competizione interspecifica su due popolazioni i e j, venne aggiunto il termine che fa sì che le equazioni di Lotka e Volterra siano:





- (eq. 1,1)
- (eq. 1,2)


In queste due equazioni il termine α12 rappresenta l'effetto che la specie 2 ha sulla popolazione della specie 1 ed α21 il viceversa (ossia l'effetto della specie 1 sulla popolazione della specie 2); α viene anche detto coefficiente di competizione.
Equilibrio e coesistenza
Affinché due specie si trovino in equilibrio esse devono avere crescita zero, ossia:


Queste condizioni si verificano solo se:


Portando a primo membro ed si avrà:
- (eq. 1,3)
- (eq. 1,4)


In assenza di competizione interspecifica, ovvero , risulterà che l'ammontare d'equilibrio di una delle due specie considerate sarà pari alla capacità portante K ( e ). Ciò significa che, prendendo solo in considerazione l'equazione 1,3, la capacità portante dell'ambiente viene ridotta della quantità dalla competizione interspecifica.
Se il termine dell'equazione 1,3 viene sostituito da si otterrà:






da cui discende che
- ,

- ,

- (eq. 1,5)

Sostituendo al termine dell'equazione 1,4 si avrà:

- (eq. 1,6)

Generalmente i coefficienti di competizione sono minori di 1 per cui il denominatore delle equazioni 1,5 ed 1,6 in genere è positivo. Affinché le due specie possano coesistere in condizioni di equilibrio si deve avere che ed siano entrambi positivi ovvero che siano positivi anche i denominatori delle due equazioni. In termini generali è necessario, per la coesistenza, che:

Le isocline zero
L'equazione 1,3 può essere riarrangiata in questo modo:

I punti della retta di quest'equazione rappresentano le condizioni in cui la crescita della popolazione sia nulla. La retta in questione viene detta isoclina zero.

Nel grafico è rappresentata l'isoclina zero per la specie 1 e si può notare che se la popolazione si trova ad un punto a sinistra della retta, tenderà ad aumentare perché è ad un valore inferiore rispetto alla capacità portante . Se, invece, la popolazione si trova su un punto a destra della retta, allora tenderà a diminuire. La retta intercetta l'asse x quando il valore è uguale alla sua capacità portante e la popolazione è uguale a zero, mentre intercetta l'asse y al valore in cui la sua capacità portante è riempita dagli individui della popolazione e nessun individuo della popolazione è presente.


Un ragionamento analogo lo si può condurre per la specie 2 utilizzando l'equazione 1,4 per tracciare l'isoclina. In questo caso se si trova ad un punto in basso rispetto alla retta la popolazione tenderà all'aumento mentre, al contrario, diminuirà qualora si dovesse trovare su un punto in alto rispetto all'isoclina.
I quattro scenari
Le due isocline zero ottenute possono essere giustapposte tra di loro in uno stesso grafico secondo modalità differenti che variano a seconda dei valori di K e di α d'entrambe le specie studiate. In tale modo si possono ottenere quattro diverse combinazioni che danno vita ad altrettanti scenari.
- e



Come si può vedere dal grafico sovrastante risulta che e che , da cui, riarraggiando, si ottengono le disuguaglianze precedentemente esposte. Il significato biologico della prima disuguaglianza risulta essere che la specie 1 () è in grado di inibire se stessa più di quanto venga limitata dalla specie 2 (). La seconda disuguaglianza, invece, significa che la specie 1 è in grado di inibire la specie 2 più di quanto essa venga limitata dalla competizione intraspecifica.
In conclusione emerge che la specie 1 è un forte competitore interspecifico mentre la specie 2 non lo è.
Come si può evincere dal grafico l'isoclina della specie 1 è completamente esterna rispetto a quella della specie 2. Se ci si trova ad un punto interno al triangolo formato dall'isoclina della specie 2 (entrambe le popolazioni sono piccole) e dagli assi cartesiani il sistema tenderà a spostarsi in alto ed a destra. Se ci si trova su in punto esterno all'isoclina della specie 1 (entrambe le popolazioni sono grandi), invece, si sposterà in basso ed a sinistra. Una volta entrato nell'area compresa tra le due isocline la traiettoria del punto si sposterà in basso ed a destra fino a raggiungere il valore (la specie 1 ha portato la specie 2 all'estinzione ed ha raggiunto la propria capacità portante.



- e



In questo caso la situazione è opposta alla precedente. La specie 2 è un forte competitore interspecifico che porta all'estinzione la specie 1 e raggiunge la propria capacità portante.
- e

In questo caso ci si trova di fronte a due bassi competitori interspecifici e vengono limitati soprattutto dalla competizione intraspecifica. Il punto d'intersezione delle due isocline corrisponde ad un punto d'equilibrio stabile cui tende il sistema. Poiché nessuna delle due specie si estingue, allora risulta possibile la coesistenza.
- e
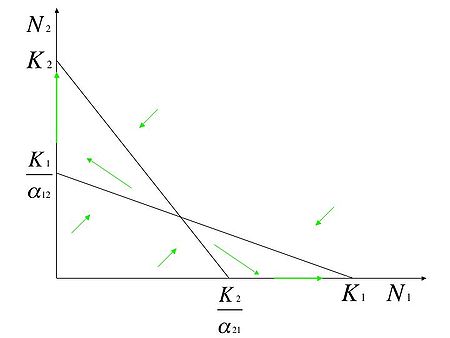
Si tratta di due popolazioni che sono inibite più dalla competizione interspecifica che da quella intraspecifica.
Il punto d'incrocio delle due isocline corrisponde ad un punto d'equilibrio instabile cui il sistema tende ad allontanarsi. Esistono, in questo caso, due punti d'equilibrio stabili corrispondenti a e . Nel primo caso la specie 2 si estingue mentre nel secondo caso è la specie 1 ad estinguersi. La specie che all'inizio detiene la più elevata densità di popolazione sarà quella che risulterà vincente.

Modello ad N specie
Il modello precedentemente esposto può essere generalizzato per ogni numero di specie tra loro competitive. L'equazione per ogni popolazione diviene:


con la sommatoria che va da 1 ad N (numero delle popolazioni).
Nel caso in cui , si avrà:


I limiti del modello
Successivi esperimenti di competizione evidenziarono delle discrepanze rispetto al modello precedentemente esposto.
Francisco Ayala (1970), ad esempio, rilevò che due specie di Drosophila (Drosophila pseudoobscura e Drosophila serrata) coesistono alla temperatura di 23 °C calcolando i valori dei coefficienti di competizione risultò che era uguale a 1,49 e aveva il valore di 3,86. Il loro prodotto era uguale a 3,86.
Avendo visto in precedenza che per avere le coesistenza deve risultare che , risulta chiaro che il valore ottenuto non è compatibile con il modello di Lotka-Volterra. Ciò indica come tale modello lineare non sia sufficiente per descrivere in maniera adeguata le dinamiche delle popolazioni in alcune situazioni.

La competizione interspecifica dipende anche da fattori ambientali
I lavori di Thomas Park (1954, 1962) permisero di dimostrare un delle ipotesi formulate da A.G. Tansley che negli studi condotti su due specie del genere Galium (Galium sylvestre e Galium saxatile) aveva espresso l'idea che il risultato della competizione interspecifica potesse dipendere anche dalle condizioni ambientali.
Effettivamente Park dimostrò che i risultati della competizione di due specie di coleotteri tenebrionidi (Tribolium castaneum e Tribolium confusum) variano a seconda della temperatura e dell'umidità ambientali. In caso di condizioni fredde ed aride prevale Tribolium castaneum mentre in condizioni calde ed umide ha la prevalenza Tribolium confusum.
Bibliografia
- Robert E. Ricklefs: Ecologia, seconda edizione italiana; Zanichelli.
- Colin R. Towsend, John L. Harper, Michael Begon: L'essenziale di ecologia; Zanichelli
Collegamenti esterni
- (EN) The struggle for existence, su ggause.com. URL consultato il 29 ottobre 2006 (archiviato dall'url originale il 28 novembre 2016).
- (EN) Iterspecific competition, su tiem.utk.edu. URL consultato il 6 luglio 2008 (archiviato dall'url originale il 9 febbraio 2008).