

Il tessuto connettivo (lat. textus conexivus) è uno dei quattro tipi fondamentali di tessuto che compongono il corpo degli animali più abbondanti nell'organismo. Questo insieme di tessuti è sempre stato definito “connettivo” impropriamente in quanto è preferibile utilizzare la definizione “tessuti trofo-meccanici”: ogni sottospecie della categoria svolge funzione trofica e/o funzione meccanica.
Descrizione generale
Viene definito come vari tipi di tessuto che hanno in comune la funzione di provvedere al collegamento, al sostegno, alla protezione, al nutrimento, alla difesa immunitaria, al trasporto di fluidi, alla riserva energetica di altri tessuti dei vari organi e che deriva dal tessuto connettivo embrionale, il mesenchima (che si origina principalmente dal mesoderma).
Istologicamente, quindi il tessuto connettivo può essere suddiviso in diversi sottotipi, a seconda delle loro prerogative morfologiche e funzionali, tutti caratterizzati dal fatto di essere costituiti da cellule non addossate le une alle altre, ma disperse in una più o meno abbondante sostanza intercellulare o matrice extracellulare costituita da una componente amorfa e da una componente fibrosa.
Cellule del tessuto connettivo
Il tessuto connettivo possiede un'ampia varietà di cellule, deputate a svolgere attività diverse in relazione anche alla natura del tessuto a cui appartengono e alla posizione che questo assume nell'organismo. In generale, è possibile operare una distinzione tra le cellule deputate alla formazione e al mantenimento della matrice (fibroblasti, condroblasti, osteoblasti, cementoblasti, odontoblasti), cellule deputate alla difesa dell'organismo (macrofagi, mastociti, leucociti) e cellule deputate a funzioni speciali, come gli adipociti del tessuto adiposo, che accumulano grassi come riserva energetica del corpo. È possibile anche distinguerle in base al loro ciclo vitale in cellule fisse (macrofagi fissi, fibroblasti, adipociti), che svolgono tutta la loro vita nel tessuto connettivo, e cellule migranti (granulociti neutrofili, linfociti, macrofagi) che invece raggiungono il tessuto connettivo dalla circolazione sanguigna. Alcune di esse, come i linfociti, possono passare liberamente dal circolo sanguigno al connettivo, altre, come i granulociti neutrofili, una volta spostatisi per diapedesi nel connettivo non possono più far ritorno nel sangue.
Fibroblasti

I fibroblasti sono le cellule più numerose del tessuto connettivo propriamente detto. La loro funzione è quella di produrre le fibre e i componenti macromolecolari della matrice extracellulare, che costituisce l'elemento di gran lunga più abbondante del tessuto, e dalla quale dipendono le funzioni di sostegno proprie del connettivo. I fibroblasti sono generalmente di aspetto fusiforme, sebbene ne esistano varietà che presentano morfologie anche diverse, come un aspetto stellato o tentacolare. Si trovano generalmente dispersi nella matrice da loro stessi creata, ed in molti casi sono disposti lungo le fibre.
Al microscopio elettronico è possibile notare, in zona perinucleare, l'apparato di Golgi e i due centrioli, i mitocondri sono generalmente lunghi e sottili, ma nei cementoblasti e negli odontoblasti assumono anche forma tondeggiante. Il reticolo endoplasmatico presenta cisterne appiattite e il suo sviluppo dipende dallo stato funzionale della cellula, tutti i filamenti del citoscheletro sono molto sviluppati, particolarmente i microfilamenti actinici concentrati nella zona corticale. Numerose le strutture di adesione quali podosomi e adesioni focali.
Quando cessano la loro attività biosintetica, i fibroblasti si trasformano in fibrociti, che hanno citoplasma debolmente acidofilo in confronto ai fibroblasti biosinteticamente attivi che lo hanno basofilo. Pertanto, fibroblasti e fibrociti rappresentano i due momenti funzionali di una stessa cellula. Cellule di funzione analoga sono presenti nei diversi sottotipi di tessuto connettivo, anche se presentano in alcuni casi peculiarità funzionali.
Corrispondenti per funzione dei fibroblasti negli altri tipi di tessuto connettivo sono:
- i condroblasti producono la matrice del tessuto cartilagineo.
- gli osteoblasti producono la matrice del tessuto osseo, caratterizzata dal fatto di essere calcificata.
- i cementoblasti e gli odontoblasti producono la matrice nei denti.
Macrofagi
I macrofagi sono, per diffusione, le seconde cellule più numerose del tessuto connettivo propriamente detto. I macrofagi sono di tipo fisso, presente nel tessuto connettivo in condizioni normali, o di tipo migrante, che si trova nel caso di danno tissutale come un processo infiammatorio. In realtà si tratta dello stesso tipo cellulare in diversa forma, per cui si preferisce adottare la distinzione in macrofago non attivato e in macrofago attivato. Al microscopio elettronico i macrofagi appaiono come cellule tondeggianti, fusiformi o stellate con un diametro di 10-30 µm, provviste di estroflessioni citoplasmatiche simili a villi.
Nel citoplasma l'apparato di Golgi e il reticolo endoplasmatico rugoso sono molto sviluppati, sono inoltre presenti numerosi lisosomi e fagosomi, un citoscheletro sviluppato con filamenti intermedi di vimentina dello spessore di 10 nm e microfilamenti actino-simili dello spessore di 6 nm, fondamentali in quanto costituiscono l'impalcatura dei "villi" del macrofago. Il nucleo è unico. Poiché a livello citoplasmatico i fibroblasti e macrofagi fissi sono piuttosto simili, per distinguerli è necessario mettere a prova la spiccata capacità fagocitaria del macrofago tramite la granulopessia, cioè l'ingestione da parte della cellula di un colorante vitale. Il vivace spostamento dei macrofagi, una volta attivati, è determinato dall'ondulazione della loro membrana plasmatica, ed è di tipo ameboide. La direzione dello spostamento è determinata da chemiotassi.
La capacità fondamentale dei macrofagi è certamente la fagocitosi, meccanismo che utilizzano con funzione difensiva. Sono infatti deputati ad assorbire ed eliminare elementi esterni, quali virus, batteri, cellule tumorali, cellule ematiche invecchiate, molecole dannose all'organismo. Il macrofago è stimolato da numerosi fattori chimici che si legano al corpo estraneo, quali per esempio gli anticorpi IgG e IgM, esso li riconosce, si attiva, ed inizia una serie di azioni volte alla sua distruzione e al coordinamento della risposta immunitaria. Per fagocitarlo, emette pseudopodi (estroflessioni della membrana plasmatica) che circondano il corpo estraneo, inglobandolo nel citoplasma del macrofago con un sistema "a cerniera". Qui è digerito dalle idrolasi acide contenute nei lisosomi e da enzimi quali il lisozima, che scinde la membrana plasmatica di molti batteri, e la mieloperossidasi.
Se il corpo estraneo è troppo voluminoso per un solo macrofago, queste cellule possono aggregarsi in complessi polinucleati (fino ad un centinaio di nuclei), chiamati cellule giganti da corpo estraneo. Contemporaneamente all'azione fagocitaria, il macrofago secerne ossido nitrico (NO) e prostaglandine, che inducono vasodilatazione, interleuchina-1 (IL-1) che attira linfociti e granulociti neutrofili, citochine, che attivano la proliferazione delle cellule circostanti, ne aumentano la capacità fagocitaria e le attirano verso la sede dell'infiammazione, eritropoietina, che stimola la maturazione dei precursori degli eritrociti nel midollo osseo, CSF (colony stimulating factors) che agiscono sulla maturazione di molte altre cellule emopoietiche. I macrofagi sono anche APC-cells (antigen presenting cells) in quanto presentano sulla loro membrana gli antigeni parzialmente elaborati dai batteri fagocitati, permettendone il riconoscimento da parte dei linfociti. Tale presentazione però è non-specifica a differenza di quella dei linfociti B.
Linfociti
I linfociti sono cellule appartenenti al sistema immunitario, e, pur essendo formalmente cellule connettivali, si trovano in prevalenza libere nel sangue. Vengono suddivisi in due classi principali: i linfociti B e i linfociti T:
- I linfociti B sono in grado di riconoscere l'antigene presentato dai macrofagi, ed in risposta maturano in plasmacellule, producendo anticorpi che intervengono ad eliminare i corpi estranei.
- I linfociti T, oltre a cooperare con i linfociti B e con le proteine del complesso maggiore di istocompatibilità per permettere il riconoscimento degli antigeni, sono anche deputati alla risposta self, ovvero all'eliminazione di cellule appartenenti all'organismo stesso, alterate dall'infezione di un virus o cancerogene.
Mastociti
I mastociti sono cellule dal diametro di 20-30 µm, hanno forma tondeggiante o fusata e sono mobili. Sono muniti di sottili estroflessioni della membrana plasmatica, discreto corredo mitocondriale, reticolo endoplasmatico e un piccolo apparato di Golgi. Il nucleo è reniforme e presenta cromatina dispersa.
La caratteristica morfologica più importante per distinguerli è la presenza, nel citoplasma, di numerosi granuli tondeggianti ed elettrondensi, omogenei nell'uomo, solubili in acqua, che si colorano metacromaticamente con i coloranti basici come il blu di toluidina, oppure con i coloranti per glicosaminoglicani solfati come l'Alcian blu. I granuli sono rivestiti da membrana e contengono eparina ed istamina. L'eparina, un glicosaminoglicano che rende ragione della colorazione di questi granuli, è un anticoagulante, mentre l'istamina è un vasodilatatore ed aumenta la permeabilità dei capillari sanguigni. La liberazione di questi granuli avviene in numerose reazioni immunologiche, in particolare in quelle di ipersensibilità immediata, cioè quando un organismo viene a contatto con un antigene per cui è già stato sensibilizzato in precedenza. È sufficiente che due recettori del mastocita (associatisi ad IgE durante la prima esposizione) vengano a contatto con l'antigene per determinarne la degranulazione.
La degranulazione consiste nello spostamento dei granuli verso la membrana plasmatica, nella fusione della membrana dei granuli con essa e nella liberazione del contenuto dei granuli nello spazio extracellulare. In tal caso si dice che avviene una degranulazione asincrona. È però possibile in casi particolari, che la risposta immunitaria si estenda a interi organi o apparati, e che la degranulazione risulti anafilattica. In questo caso i granuli si fondono tra loro e vengono espulsi violentemente dal mastocita determinando lo shock anafilattico. Un mastocita è in grado di ricostituire il proprio corredo granulare in 1-2 giorni dalla degranulazione. Sono anche in grado di secernere sostanze quali le interleuchine 4,5,6 (IL-4, IL-5, IL-6), citochine e fattori chemiotattici. L'attivazione dei mastociti comporta la liberazione di leucotrieni, che vengono sintetizzati a partire dall'acido arachidonico contenuto in alcuni piccoli granuli lipidici del citoplasma della cellula. I leucotrieni inducono la contrazione della muscolatura liscia delle vie aeree e sono coinvolti nelle crisi asmatiche.
Adipociti
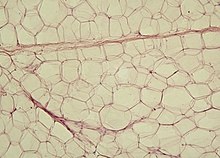
Gli adipociti sono cellule fisse del tessuto connettivo adibite alla raccolta, al mantenimento e alla secrezione dei lipidi. Hanno un diametro molto variabile, che può superare i 100 µm forma tondeggiante a causa dell'unica goccia lipidica (per gli adipociti uniloculari) che occupa quasi tutto il citoplasma, schiacciando il nucleo contro la membrana plasmatica. Si trovano in tutti i tipi di tessuto connettivo, lungo i vasi sanguigni, e costituiscono il tipo cellulare prevalente nel tessuto adiposo. Svolgono una funzione di riserva energetica, contribuendo al riscaldamento del corpo, oltre a produrre ormoni(ormoni steroidi) e fattori di crescita. Possono essere colorati con i coloranti solubili nei grassi come Sudan Nero, Sudan III o Orange G. Gli adipociti esistono in tre varietà: gli adipociti uniloculari, gli adipociti multiloculari e gli adipociti pauciloculari.
- gli adipociti univacuolari (Uniloculari) presentano un unico grande vacuolo, contenente i lipidi, che riempie la quasi totalità della cellula. Il nucleo ed il citoplasma cellulare risultano perciò decentrati e schiacciati lungo i bordi della membrana plasmatica. Essi formano il tessuto adiposo bianco.
- gli adipociti multivacuolari (Multiloculari) non possiedono invece il vacuolo centrale, ma presentano i lipidi raccolti in numerose piccole gocce disperse nel citoplasma. In queste cellule il nucleo si presenta in posizione centrale. Formano il tessuto adiposo bruno.
Matrice extracellulare
Tutte le cellule dei diversi tipi di tessuto connettivo si trovano disperse in una sostanza gelatinosa, liquida o solida denominata matrice o matrice extracellulare. La matrice extracellulare è costituita da una porzione fibrosa, composta da proteine, inclusa in una soluzione acquosa di proteine, glicoproteine e proteoglicani. Le proteine in questione sono: collagene, elastina, laminina, fibronectina, condronectina e osteonectina\SPARC.
La matrice extracellulare è quindi divisibile in:
- una matrice di materiale amorfo detta sostanza fondamentale;
- una componente fibrillare.
Da un punto di vista istologico, al microscopio la componente amorfa viene allontanata durante i processi di allestimento: accanto alla componente fibrosa si trovano delle lacune bianche che in vivo sono occupate dalla sostanza amorfa.
Fibre
Le fibre del tessuto connettivo sono immerse nella sostanza amorfa, e conferiscono stabilità strutturale alla matrice. Le fibre sono distinte in tre tipi fondamentali, a seconda della loro composizione e struttura:
- fibre collagene
- fibre reticolari
- fibre elastiche
Le fibre collagene e le fibre reticolari sono costituite entrambe da molecole di procollagene, ma differiscono tra loro per l'organizzazione spaziale di tali molecole; le fibre elastiche sono invece costituite da due catene proteiche di diversa natura: la fibrillina e l'elastina.
Fibre collagene
Le fibre collagene sono il tipo di fibre più rappresentato dell'organismo umano e nei tessuti connettivi, rappresentano da sole il componente non minerale più abbondante dopo l'acqua, costituendo fino al 6% del peso corporeo. Appaiono come lunghe fibre bianche ondulate, che si diramano in più direzioni (nel caso di un tessuto connettivo denso irregolare o connettivo lasso) o in un'unica direzione (tessuto connettivo denso regolare), hanno uno spessore variabile da 1 a 12 µm. Ogni fibra collagene è costituita da decine di fibrille più sottili, del diametro di 0,2-0,3 µm, che determinano la sua striatura longitudinale, immerse in una sostanza amorfa.
Ogni fibrilla collagene è a sua volta costituita da microfibrille che si associano longitudinalmente tra loro, determinandone la birifrangenza. Le microfibrille, esaminate al microscopio elettronico, appaiono striate trasversalmente al loro asse maggiore, in particolare le striature si ripetono ogni 70 nm a fresco oppure ogni 64 nm a secco, si dice perciò che possiedono una periodicità assile di 64-70 nm. Sono distinguibili due tipi di striature trasversali, una più elettrondensa e l'altra meno elettrondensa. Dal momento che le molecole di tropocollagene si associano tra loro in maniera sfasata, sovrapponendosi per un quarto della loro lunghezza, si possono spiegare i due tipi di striature affermando che le bande meno elettrondense sono costituite dalle teste delle molecole di tropocollagene e dall'estremità delle code, mentre le bande più elettrondense sono costituite tra le code delle molecole di tropocollagene e gli intervalli tra una molecola e la successiva.
Le fibre collagene sono molto resistenti alla trazione, flessibili, ma praticamente inestensibili. In soluzione acida diluita esse tendono a gonfiarsi, mentre sono disciolte in soluzioni contenenti acidi o basi forti, oltre ad essere digerite specificamente dall'enzima collagenasi. La denaturazione del collagene, che può avvenire per bollitura, porta le fibre a trasformarsi in una sostanza gelatinosa. Il collagene è sintetizzato principalmente da fibroblasti, condroblasti e osteoblasti, ma può essere prodotto anche dalle cellule epiteliali, com'è il caso del collagene di tipo IV, che forma la lamina basale. Le fibre collagene sono evidenziate in microscopia ottica attraverso coloranti acidi come il blu di anilina nella tecnica di colorazione Azan-Mallory, assumono l'eosina e sono PAS-negative o leggermente PAS-positive a causa di brevi catene laterali carboidratiche costituite da galattosio o glucosil-galattosio legate alle molecole di idrossilisina. Esistono 25 tipi diversi di catene α che si associano tra loro in triplette (una molecola di tropocollagene è costituita da tre α-eliche) in modo da formare 29 diversi tipi di collagene. I 29 tipi di collagene sono divisibili in tre classi, le quali sono:
- Collageni fibrillari: sono le fibre collagene più comuni, da sole costituiscono la quasi totalità del collagene nel corpo umano, vi appartengono i collageni di tipo I, II, III e V. Il collagene di tipo I costituisce il 90% del collagene nel corpo, costituisce le ossa, i tendini, le fibre collagene del derma e la dentina. Il collagene di tipo II è diffuso nella cartilagine e nell'umor vitreo. Il collagene di tipo III è diffuso nel derma, nei muscoli e nella parete dei vasi sanguigni. Il collagene di tipo V è diffuso nel tessuto interstiziale.
- Collageni associati a fibrille: sono fibre collagene che non si riscontrano mai da sole, ma si associano sempre ai collageni fibrillari all'interno delle loro fibrille o formano legami tra le fibrille e la matrice circostante. Vi appartengono i collageni di tipo IX e XII, il primo si associa al collagene di tipo II nella cartilagine, il secondo si associa al tipo I e III nel derma e nei tendini.
- Collageni laminari o reticolari: sono fibre collagene che non si organizzano in spessi fasci ma in maglie reticolate, spesso localizzate negli spazi pericellulari o nella membrana basale. Ne fanno parte i collageni di tipo IV, che costituisce gran parte della membrana basale, VIII che si associa agli endoteli, e X presente nelle cartilagini di coniugazione delle ossa.
Fibre reticolari
Le fibre reticolari, costituite da catene di collagene di tipo III, sono diffuse nel tessuto connettivo lasso, nei muscoli, nell'endonevrio, nel tessuto adiposo, negli organi linfoidi e nella parete dei vasi sanguigni. Sono anch'esse costituite da fibrille e microfibrille che presentano periodicità assile di 64-70 nm, ma le fibrille sono più sottili (spessore medio di 50 nm) e di conseguenza lo sono anche le fibre reticolari (spessore variabile 0,5-2 µm). Le fibre reticolari non si associano tra di loro per formare fasci, ma costituiscono trame e reti sottili, decorrendo su due piani o in senso tridimensionale, con ampi spazi tra le maglie occupati da matrice amorfa. Non possiedono la striatura longitudinale delle fibre collagene, ma possiedono un maggiore grado di glicosilazione dell'idrossilisina e per questa ragione sono PAS-positive, inoltre si colorano facilmente con il metodo dell'impregnazione argentica e per questo sono anche definite fibre argirofile.
Fibre elastiche

Le fibre elastiche o fibre gialle sono una componente dei tessuti connettivi in alcune strutture del corpo. Esse sono fondamentali, ad esempio, nella funzionalità delle arterie, in particolare per l'aorta. Le fibre elastiche si trovano nella matrice extracellulare insieme alle fibre collagene e sono formate da proteine quali l'elastina e la fibrillina. Il tessuto elastico rientra nella categoria dei connettivi propriamente detti. Esse vengono colorate attraverso l'orceina, che fa assumere loro una caratteristica colorazione marrone, oppure attraverso il metodo fucsina-resorcina di Weigert.
Sostanza amorfa
La sostanza amorfa (o sostanza fondamentale o anista) costituisce un gel compatto nel quale sono immerse le fibre. È costituita essenzialmente da macromolecole di origine glucidica chiamate glicosaminoglicani (GAG) e da associazioni di questi ultimi con proteine, definite proteoglicani.
- I glicosaminoglicani sono i più importanti ed abbondanti componenti della matrice amorfa. Si tratta di lunghi polimeri, con massa atomica variabile da poche migliaia a milioni di Da, costituiti da catene di disaccaridi ripetuti decine di volte, a loro volta formati da un acido uronico (D-glucoronato, L-iduronato) legato ad un amino-zucchero (N-acetil-D-glucosamina, N-acetil-D-galattosamina). I glicosaminoglicani possono essere solforati (cheratan-solfato, condroitin-solfato, eparan-solfato, dermatan-solfato, eparina) oppure non solforati (acido ialuronico). Il glicosaminoglicano più importante è l'acido ialuronico, che costituisce anche la catena centrale degli aggregati proteoglicanici. I glicosaminoglicani riescono a legare notevoli quantità di acqua.
- I proteoglicani sono costituiti da numerosi glicosaminoglicani associati trasversalmente ad una proteina che funge da catena centrale, è in questo stato che si trova la maggior parte dei glicosaminoglicani della matrice, fatta eccezione per l'acido ialuronico che, per la sua elevata viscosità non si lega, contribuendo a formare, tra le altre cose, il liquido sinoviale. Il peso molecolare di un proteoglicano varia da 1 a 10 milioni di dalton, di cui l'80-95% è composto da glicosaminoglicani e il 5-20% da proteine. Sono sintetizzati nell'apparato di Golgi che lega uno specifico tetrasaccaride (xilosio-galattosio-galattosio-acido glucuronico) ai residui di serina della proteina centrale, per poi aggiungere un monosaccaride alla volta all'estremità libera del tetrasaccaride. Alcuni dei proteoglicani più importanti sono l'aggrecano, presente nella matrice cartilaginea, il sindecano, il versicano, il neurocano, la decorina e il β-glicano. I proteoglicani possono inoltre unirsi intorno ad una molecola di acido ialuronico centrale, formando delle strutture di ordine superiore definite aggregati (o complessi) proteglicanici, che sono fra le molecole organiche più grandi esistenti in natura, con peso di decine milioni di Da per diversi μm di lunghezza, dimensioni paragonabili a quelle di un batterio. I proteoglicani a causa della loro struttura, della viscosità e della permeabilità costituiscono degli ottimi filtri molecolari che possono diffondere alcune sostanze di basso peso molecolare, intrappolarne altre più voluminose, impediscono l'attacco di cellule del sangue a causa della loro carica negativa, possono fungere da recettori sulla membrana plasmatica o più comunemente nel glicocalice.
- Le glicoproteine, in quantità inferiori alle due categorie precedenti, tra le quali spicca la fibronectina, che, grazie all'interazione stabilizzatrice dei glicosaminoglicani solfati, si lega alle fibre collagene.
A causa della scarsa densità delle macromolecole che la costituiscono, è la sostanza amorfa trasparente ed invisibile al microscopio a fresco. È leggermente PAS-positiva per il suo contenuto in glicoproteine (è intensamente PAS-positiva nella cartilagine, nelle membrane basali e nell'osso dove la concentrazione di glicoproteine è maggiore), può però essere colorata con il metodo Alcian blu e con i coloranti basici come l'anilina, che danno luogo a fenomeni di metacromasia. La metacromasia è dovuta alla presenza dei glicosaminoglicani acidi della matrice, ed è tanto più elevata quanto più questi sono solforati (condroitin-solfato, cheratan-solfato, eparan-solfato). La sostanza amorfa contiene elevate quantità d'acqua, le cui molecole (H2O) però difficilmente si presentano come liquido interstiziale o tissutale libero, ma sono legate alle molecole della matrice, determinandone l'idratazione. L'acqua legata alla matrice, in cui sono disciolti gas ed altre sostanze, diffonde a partire dai capillari sanguigni e funziona come mezzo di dispersione e scambio tra la circolazione sanguigna e il tessuto connettivo, permettendone il nutrimento. Si dice quindi che la sostanza amorfa sia l'elemento con funzione trofica del tessuto connettivo. È possibile riscontrare nella matrice amorfa grandi quantità di liquido interstiziale libero in caso di infiammazione. Oltre alla sua funzione trofica, la disposizione delle molecole della matrice influenza l'orientamento delle fibre in essa contenute, e con la sua complessa trama ostacola la diffusione di microrganismi ed agenti patogeni.
Tipi di tessuto connettivo

Tessuto connettivo
Vi sono diversi tipi di tessuto connettivo, classificati in base a criteri morfologici e funzionali. Il tessuto connettivo più comune, a cui ci si riferisce in genere utilizzando questo termine, viene definito tessuto connettivo propriamente detto (abbreviato spesso come “tessuto connettivo p.d.”). Esso svolge funzioni di sostegno e di protezione, costituisce la base su cui poggiano i diversi epiteli e contribuisce alla difesa dell'organismo contro urti e traumi esterni.
Il tessuto connettivo propriamente detto si suddivide in:
Tessuto connettivo denso
Si distingue per l'abbondanza della componente fibrosa raccolta in fasci, rispetto alla sostanza amorfa e alla componente cellulare. Per il tipo di fibre che lo compongono può essere ulteriormente suddiviso in fibroso (fibre collagene tipo I) o elastico (fibre elastiche), e per la disposizione delle fibre può essere distinto in regolare, se esse assumono andamento ordinato, o irregolare se non assumono una disposizione ordinata. La funzione del tessuto connettivo denso è prevalentemente meccanica, l'orientamento e la qualità delle sue fibre determinano infatti le sue diverse proprietà, come la resistenza alla trazione o la deformabilità.
- Tessuto connettivo denso irregolare: è un connettivo caratterizzato da numerose fibre collagene che si aggregano in fasci molto densi fra loro, talvolta accompagnati da reti di tessuto elastico. Le cellule sono poche, vi sono perlopiù fibroblasti e rari macrofagi, scarsa la sostanza amorfa. Si riscontra nel derma, nella capsula fibrosa di organi quali milza, fegato, testicolo, linfonodi, forma la guaina dei tendini e dei nervi più importanti e il periostio.
- Tessuto connettivo denso regolare: è un connettivo caratterizzato da fibre collagene fittamente stipate ed orientate tutte nella stessa direzione, concorde a quella della trazione che il tessuto deve sopportare. Scarsa la sostanza amorfa, pochissime le cellule, che sono quasi esclusivamente fibroblasti disposti nei sottili interstizi delle fibre collagene. Come nel denso irregolare, alle fibre collagene possono essere associate reti di tessuto elastico. Forma tendini, legamenti, aponeurosi, stroma corneale. Ivi tra i fasci paralleli di fibre collagene vanno inserendosi fibroblasti specializzati chiamati tendinociti o tenociti, dei quali si può apprezzare in una visione trasversale la peculiare forma citoplasmatica stellata, ma i cui prolungamenti vanno confondendosi con le fibre collagene se osservati in seguito ad un taglio longitudinale. La sostanza del tendine è circondata da un connettivo poco ordinato detto epitendineum. Nei tendini e nei legamenti le fibre raggiungono la disposizione più ordinata e sono orientate tutte nella stessa direzione con i fasci legati da tessuto connettivo lasso, nelle aponeurosi le fibre sono disposte in strati ordinati in più direzioni, nello stroma corneale invece questi strati sono orientati perpendicolarmente l'uno all'altro.
- Tessuto connettivo denso elastico: è un connettivo caratterizzato dalla prevalenza delle fibre elastiche sulle fibre collagene, vi sono fibroblasti interposti tra i fasci di fibre elastiche, a loro volta avvolti da fibre reticolari. Forma i legamenti gialli delle vertebre, le corde vocali, le lamine fenestrate delle arterie maggiori.
Tessuto connettivo lasso
È il tessuto connettivo propriamente detto più diffuso. Si distingue per l'abbondanza della sostanza amorfa rispetto alla componente fibrosa e su quella cellulare e per il maggior numero di nuclei cellulari rispetto al connettivo denso. Per il tipo di fibre che lo compongono può essere ulteriormente classificato come:
- fibroso (fibre collagene tipo I),
- reticolare (fibre collagene di tipo III),
- elastico (fibre elastiche).
Il tessuto connettivo lasso reticolare è particolarmente diffuso negli organi emopoietici e linfoidi, nella muscolatura liscia e in alcune ghiandole; tra le sue fibre sono presenti numerosi macrofagi e fibroblasti. Uno speciale tipo di tessuto connettivo lasso è il tessuto mucoso, diffuso nell'embrione e in particolare costituente della "gelatina di Wharton", ovvero la sostanza amorfa del funicolo ombelicale. Tale tessuto si definisce mucoso a causa della sua consistenza, dovuta all'abbondante quantità di acido ialuronico. Possiede poche fibre collagene o reticolari, scarsi macrofagi ma numerosi fibroblasti stellati. Se colorato, presenta intensa basofilia. Il tessuto connettivo lasso forma la tonaca propria e la tonaca sottomucosa delle mucose, avvolge molti organi e si inoltra in essi con setti che ne suddividono il parenchima in lobi e lobuli, costituisce inoltre lo stroma, la tonaca intima e la tonaca avventizia delle arterie, la tonaca media e avventizia delle vene assieme al tessuto muscolare liscio. Connette gli organi e ne riempie gli spazi liberi, circonda muscoli (epimisio, perimisio) e nervi (endonevrio, perinevrio).
Tessuto adiposo
Il tessuto adiposo, che più propriamente andrebbe chiamato organo adiposo, è un particolare tipo di tessuto connettivo. Ha un colorito giallo ed una consistenza molliccia, ed è costituito da cellule adipose, dette adipociti, che possono essere singole o riunite in gruppi nel contesto del tessuto connettivo fibrillare lasso. Se le cellule adipose sono molte, e per questo sono organizzate in lobuli, allora costituiscono il tessuto adiposo che è una varietà di tessuto connettivo lasso. Questo tessuto è presente in molte parti del corpo e, in particolare, sotto alla pelle, venendo a costituire il pannicolo adiposo (lat. panniculus diminutivo di pannus, cioè panno) cioè striscia o strato di tessuto di grasso sottocutaneo particolarmente abbondante.
Per il 50% è accumulato nel tessuto connettivo sottocutaneo dove svolge sia un'azione di copertura, che un'azione meccanica che un'azione coibente. Il 45% lo ritroviamo nella cavità addominale dove forma il tessuto adiposo interno. Il 5% lo ritroviamo nel tessuto muscolare come grasso di infiltrazione che ha la funzione di agevolare e facilitare la funzione del tessuto muscolare. Questo sottotipo di tessuto è costituito dalle cellule adipose multiloculari (al contrario dei normali adipociti non hanno un'unica goccia lipidica ma tante piccole gocce che aumentano la superficie di combustibile esposta al citosol e lo rendono quindi più disponibile per il metabolismo cellulare), è molto scarso nell'uomo adulto e appare brunastro se osservato al microscopio ottico, sia per la presenza massiccia di mitocondri che per l'elevata vascolarizzazione.
Il tessuto adiposo bruno ha esclusivamente la funzione di produrre calore perché i mitocondri delle cellule adipose multiloculari hanno meno ATP sintetasi, l'enzima che catalizza la sintesi dell'ATP, a partire dall'ADP, da fosforo inorganico e dall'energia derivante dalla respirazione cellulare. Posseggono invece una proteina canale (la termogenina) la quale dissipa il gradiente elettrochimico degli ioni idrogeno che la catena di trasporto degli elettroni normalmente produce a cavallo tra la membrana interna e lo spazio intermembrana. Questa peculiarità fa sì che l'energia prodotta dalla scissione dei trigliceridi non venga utilizzata per la produzione di ATP e venga trasformata in calore.
Il grasso bruno è ben rappresentato nei neonati di molte specie (nella specie umana soprattutto a livello della nuca, del collo e delle scapole). Negli adulti è abbondante invece quasi esclusivamente nelle specie che vanno in letargo, mentre negli adulti di altre specie, compresa quella umana, esso è scarsamente presente (l'esistenza di due diversi tipi di lipoma, cioè di neoplasie del tessuto adiposo, mostra però la permanenza di due diversi tipi di tessuto adiposo anche nell'individuo adulto). (of)
Tessuto cartilagineo
Il tessuto cartilagineo è un particolare tipo di tessuto connettivo. È costituito da fibre connettivali immerse in una sostanza amorfa molto consistente chiamata condrina e di cellule contenute in cavità lenticolari. Le cellule sono disposte in gruppi isogeni e vengono chiamate condrociti. Questo tipo di tessuto si divide in: ialino, elastico e fibroso. La Cartilagine Ialina è il tipo di Cartilagine di gran lunga più diffuso nel corpo, essa è ricca di fibre collagene immerse in una matrice "gommosa" dall'aspetto vitreo (ialino significa appunto "vitreo"). La Cartilagine Ialina ricopre le estremità, forma parte del naso, lo scheletro della laringe e dei bronchi, e congiunge le costole allo sterno. Mentre la Cartilagine Elastica è una varietà del tessuto cartilagineo ed è quella che forma il padiglione auricolare.
Tessuto osseo
Il tessuto osseo è un tipo particolare di tessuto connettivo, che svolge funzione di sostegno strutturale dell'intero organismo. La sua caratteristica principale è quella di possedere una matrice extracellulare calcificata, che fornisce al tessuto stesso notevoli doti di compattezza e di resistenza. La matrice contiene inoltre fibre, specialmente collagene, che conferiscono al tessuto un certo grado di flessibilità, e ovviamente da cellule denominate osteoblasti. In base all'organizzazione della matrice, il tessuto osseo può essere diviso in due sottotipi: tessuto osseo lamellare e tessuto osseo non lamellare.
- il tessuto osseo non lamellare, è presente nei volatili, mentre nei mammiferi rappresenta la versione immatura del tessuto osseo, ed è presente solo durante lo sviluppo dell'organismo, per essere poi rimpiazzato dal tessuto lamellare durante la crescita. In questo tipo di tessuto la matrice calcificata non è organizzata in strutture definite, ma si presenta disordinata ed irregolare
- il tessuto osseo lamellare è presente invece nell'organismo adulto, e caratterizzato dall'alto grado di organizzazione dei componenti della matrice, che sono disposte in strati, definiti appunto lamelle, altamente ordinati. Può a sua volta essere suddiviso in due tipi, a seconda del tipo di organizzazione delle lamelle: tessuto osseo spugnoso e tessuto osseo compatto.
- nel tessuto osseo spugnoso, le lamelle vanno a costituire strutture ramificate definite spicole; per questo motivo, all'esame ottico appare come una massa spugnosa ricca di cavità intercomunicanti
- nel tessuto osseo compatto invece, le lamelle si organizzano a formare strutture concentriche, definite osteoni, addossate le une alle altre a lasciare un'unica lacuna centrale.
Tessuto ematico o sangue
Il sangue è un tessuto fluido contenuto nei vasi sanguigni dei Vertebrati, dalla composizione complessa, può essere considerato come una varietà di tessuto connettivo. È formato da una parte liquida detta siero e da una parte corpuscolare, costituita da cellule o frammenti di cellule. Ha una funzione trofica (cioè portano sostanze nutritive, ossigeno, ormoni, ecc.)
Linfa
La linfa è un altro tessuto fluido, che circola nel sistema linfatico. Si distingue dal sangue sia per la composizione molecolare del plasma, sia per il contenuto cellulare: nella linfa sono infatti del tutto assenti i globuli rossi e sono preponderanti i linfociti.
Bibliografia
- S. Adamo, P. Carinci, M. Molinaro, G. Siracusa, M. Stefanini, E. Ziparo (a cura di), Istologia di V. Monesi. Piccin Editore, 5ª edizione, ISBN 88-299-1639-0
- Don W. Fawcett, Bloom & Fawcett Trattato di Istologia. McGraw-Hill, 12ª edizione, ISBN 88-386-2050-4
- B. Young, J.W. Heath, Atlante di istologia e anatomia microscopica del Wheater. Casa Editrice Ambrosiana, 3ª edizione, ISBN 88-408-1171-0
Voci correlate
- Istologia
- Linfa (zoologia)
- Sangue
- Tessuto (biologia)
- Tessuto epiteliale
- Tessuto muscolare
- Tessuto nervoso
- Tessuto adiposo
- Tessuto cartilagineo
- Tessuto ghiandolare
- Tessuto osseo
Collegamenti esterni
- (EN) Tessuto connettivo, su Enciclopedia Britannica, Encyclopædia Britannica, Inc.
| Controllo di autorità | Thesaurus BNCF 11717 · LCCN (EN) sh85031183 · GND (DE) 4006725-7 · BNF (FR) cb119348626 (data) · J9U (EN, HE) 987007555421205171 · NDL (EN, JA) 00565679 |
|---|