
I recettori accoppiati alle proteine G (o GPCR) sono una famiglia di recettori biologici che comprende i recettori colinergici muscarinici, i recettori adrenergici, dopaminergici, serotoninergici, i recettori cannabinoidi e i recettori degli oppiacei e delle purine.
Struttura molecolare
I GPCR sono costituiti da una singola catena polipeptidica formata anche da 1100 residui. La caratteristica strutturale è rappresentata da 7 -eliche transmembrana, simili a quelle che si trovano nei canali ionici, con un dominio extracellulare N-terminale di lunghezza variabile e un dominio intracellulare C-terminale.

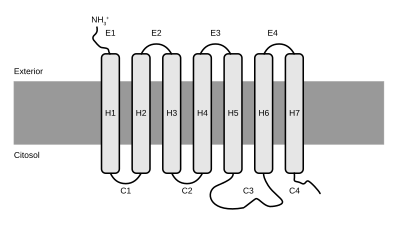
I GPCR vengono divisi in tre distinte famiglie che condividono la stessa struttura eptaelicale ma differiscono per vari aspetti, principalmente per la lunghezza della sequenza N-terminale e la localizzazione del sito di legame per l'agonista.
- La famiglia A, a cui appartiene la rodopsina, è di gran lunga la più numerosa e comprende la maggior parte dei recettori per le monoamine e i neuropeptidi.
- La famiglia B è costituita dai recettori della secretina, del glucagone e della calcitonina.
- La famiglia C è costituita principalmente dai recettori metabotropici del glutammato e dai recettori sensibili al .

Il terzo lungo loop citoplasmatico dei recettori corrisponde alla regione della molecola che si accoppia alla proteina G. Modifiche di questa porzione della proteina determinano la formazione di recettori ancora in grado di legare i propri ligandi, ma incapaci di accoppiarsi alle proteine G e di determinare i conseguenti effetti.
Grazie al miglioramento delle tecnologie di cristallografia e NMR, molte strutture vennero via via risolte a partire dal 2000, consentendo la acquisizione di conoscenze più precise per lo structure-based drug design.
Trasduzione del segnale da parte dei GPCR
Attraverso l'attivazione di vari meccanismi di trasduzione del segnale, i GPCR controllano diversi aspetti della funzione cellulare. Il collegamento tra il recettore e il primo stadio della trasduzione del segnale viene stabilito attraverso le proteine G.
Le proteine G rappresentano il livello intermedio nella gerarchia organizzativa di gestione della comunicazione tra recettori ed enzimi effettori o canali ionici. Le proteine G eterotrimeriche consistono di tre subunità , e . I nucleotidi guaninici si legano alla subunità , che è provvista di attività enzimatica, catalizzando la conversione del GTP in GDP. Le subunità e rimangono associate a formare un unico complesso . Tutte e tre le subunità sono ancorate alla membrana plasmatica mediante la catena di un acido grasso, legata alle proteine G per mezzo di una reazione chiamata prenilazione.




Allo stato di riposo, la proteina G si trova libera nella forma di trimero e con il GDP legato al sito specifico della subunità . L'occupazione di un GPCR da parte di una molecola di agonista attiva un cambiamento conformazionale, che coinvolge il dominio citoplasmatico del recettore con l'acquisizione di uno stato di alta affinità per il trimero . L'associazione del trimero con il recettore determina il rilascio del GDP legato e la sua sostituzione con il GTP; questa modificazione, a sua volta causa la dissociazione del trimero dalla proteina G con il rilascio di -GTP e delle subunità . Queste sono le forme attive della proteina G, che diffondono nella membrana e possono legarsi con enzimi e canali ionici, causandone, a seconda dei casi, l'attivazione o l'inattivazione. Il processo termina con l'idrolisi del GTP a GDP da parte della subunità , che possiede attività GTP-asica. La -GDP che così si forma, si dissocia dall'effettore e si combina con , completando in tal modo il ciclo. Poiché l'idrolisi del GTP è il passaggio che pone fine alla capacità della subunità di determinare il suo effetto, la regolazione della sua attività GTP-asica da parte dell'effettore implica che l'attivazione di quest'ultimo tende ad essere autolimitante. Il meccanismo porta a un'amplificazione del segnale, in quanto un singolo complesso agonista-recettore può attivare parecchie proteine G per volta, e ognuna di queste può rimanere associata con l'enzima effettore per tempi sufficientemente lunghi da determinare la formazione di molte molecole di prodotto. Quest'ultimo è solitamente un "secondo messaggero", per cui si verifica un'ulteriore amplificazione prima che sia evidente la risposta cellulare finale che può essere una risposta autoimmune. I ricercatori del Moores Cancer Center dell'Università della California a San Diego, hanno scoperto nel 2009 che le proteine G di comunicazione sono strettamente dipendenti dal colecalciferolo; una grave carenza di questa vitamina (Vit D3)determina una mancanza della comunicazione cellulare, e questa mancanza di comunicazione determina una regressione delle cellule a livello di staminali, in pratica sarebbe alla base di numerosi tipi di cancro; il modello è stato chiamato DINOMIT, ed ha ricevuto numerose conferme. Questa scoperta è stata pubblicata su “Le Scienze” il 22/05/2009 .

Ci sono differenze molecolari tra le varie proteine G: queste differenze danno origine a tre principali classi di proteine (, e ), che sono selettive sia per i recettori che per gli effettori con i quali si accoppiano. Le proteine e promuovono rispettivamente la stimolazione e l'inibizione dell'enzima adenil ciclasi, e un simile controllo bidirezionale è attivo su altri effettori, come la fosfolipasi C.



Bersagli per le proteine G
I principali bersagli delle proteine G, attraverso i quali i GPCR controllano diversi aspetti delle funzioni cellulari, sono i seguenti:
- adenilato ciclasi: l'enzima responsabile della formazione del cAMP;
- fosfolipasi C: l'enzima responsabile della formazione dell'inositolo trifosfato e del diacilglicerolo;
- canali ionici: in particolare i canali del calcio e del potassio.
Sistema dell'adenil ciclasi/AMPciclico
Il cAMP è un nucleotide sintetizzato all'interno della cellula a partire da ATP e con l'intervento di un enzima legato alla membrana, l'adenil ciclasi. Il cAMP viene prodotto continuamente e inattivato per idrolisi a 5'-AMP attraverso l'azione di una famiglia di enzimi noti come fosfodiesterasi. Molti farmaci, ormoni e neurotrasmettitori agiscono sui GPCR e producono i loro effetti aumentando o diminuendo l'azione catalitica dell'adenil ciclasi, determinando in tal modo un incremento o una riduzione della concentrazione intracellulare di cAMP.
Gli effetti regolatori del cAMP sulle funzioni cellulari sono molteplici e comprendono, ad esempio, enzimi coinvolti nel metabolismo energetico, nella divisione cellulare e nella differenziazione cellulare; il trasporto ionico; canali ionici e le proteine contrattili della muscolatura liscia. Tutti questi diversi effetti sono, però, provocati da un unico meccanismo, e precisamente l'attivazione di protein-chinasi da parte dell'cAMP. Le protein-chinasi determinano una regolazione funzionale di molte proteine cellulari attraverso la fosforilazione dei loro residui serinici e treoninici, utilizzando ATP come fonte di gruppi fosfato. La fosforilazione può attivare o inibire gli enzimi bersaglio o i canali ionici.
Esempi di regolazione da parte della protein-chinasi dipendente dall'cAMP includono l'aumento di attività dei canali del calcio attivati dal voltaggio nelle cellule muscolari cardiache; la fosforilazione di questi canali aumenta la quantità di calcio che entra nelle cellule durante il potenziale d'azione, incrementando in tal modo la forza di contrazione del cuore.
Nella muscolatura liscia, la protein-chinasi cAMP-dipendente fosforila, inattivandola, un'altra chinasi, la chinasi della catena leggera della miosina, che è necessaria per la contrazione. Questo spiega il rilassamento del muscolo liscio indotto da molti farmaci che aumentano la produzione di cAMP in questo sito.
Come è stato precedentemente detto, i recettori accoppiati a proteina inibiscono l'adenil ciclasi e pertanto riducono la formazione di cAMP. Esempi comprendono il recettore colinergico muscarinico (ad esempio il recettore del muscolo cardiaco), il recettore adrenergico presente nella muscolatura liscia e i recettori per gli oppiacei.


Il cAMP viene idrolizzato, all'interno delle cellule, dalla fosfodiesterasi, un enzima che è inibito da farmaci come le metilxantine (ad esempio teofillina e caffeina). La somiglianza tra alcune delle azioni di questi farmaci e quelle delle catecolamine riflette probabilmente la loro proprietà comune di aumentare la concentrazione intracellulare di cAMP.
Il sistema fosfolipasi C/inositolo fosfato
Molti ormoni, oltre a promuovere l'aumento della concentrazione di calcio libero intracellulare (ad esempio gli agonisti muscarinici e gli agonisti [[-adrenergici]] che agiscono sulla muscolatura liscia e sulle ghiandole salivari, e l'ormone antidiuretico attivo sulle cellule epatiche), aumentano anche il turnover di inositolo fosfato (PI). In questo processo gioca un ruolo chiave un membro della famiglia dei PI, e precisamente il fosfatidilinositolo 4,5-bisfosfato (), che possiede gruppi fosfato addizionali attaccati all'anello dell'inositolo. Il è un substrato dell'enzima di membrana , che lo scinde in diacilglicerolo (DAG) e inositolo 1,4,5-trisfosfato (): entrambi funzionano come secondi messaggeri. Dopo la scissione di , viene ripristinata la condizione basale. Il DAG viene fosforilato a formare l'acido fosfatidico, mentre l' viene progressivamente defosforilato e poi riaccoppiato con l'acido fosfatidico a riformare .



Fosfoinositoli e calcio intracellulare
è un mediatore solubile che viene liberato nel citosol dove si lega al suo recettore specifico (recettore ), che è un canale del calcio attivato dal ligando localizzato sulla membrana del reticolo endoplasmatico. Il ruolo principale di è quello di controllare il rilascio di dai siti di accumulo intracellulare.
Diacilglicerolo e protein-chinasi C
Il DAG e l' vengono prodotti dall'idrolisi di PI promossa dall'attivazione del recettore. L'azione principale del DAG consiste nell'attivazione di una protein-chinasi di membrana, la protein-chinasi C (PKC) che catalizza la fosforilazione di varie proteine intracellulari. Il DAG, diversamente dai fosfoinositoli, è altamente lipofilo e rimane all'interno della membrana. Si lega a un sito specifico localizzato sulla PKC, che migra dal citosol alla membrana cellulare in presenza di DAG e viene in questo modo attivata. Esistono almeno 12 sottotipi di PKC, distribuiti eterogeneamente in cellule diverse e fosforilano proteine diverse. La maggior parte viene attivata da DAG e aumenta i livelli intracellulari di .
Le varie isoforme di PKC agiscono su varie proteine funzionali come i canali ionici, i recettori, gli enzimi (tra cui altre chinasi) e alcune proteine del citoscheletro. Le chinasi, in generale, hanno un ruolo centrale nella trasduzione del segnale e controllano molti aspetti della funzione cellulare.
Canali ionici come bersaglio delle proteine G
I GPCR possono controllare l'attività funzionale di canali ionici direttamente, attraverso meccanismi che non implicano il coinvolgimento di secondi messaggeri come il cAMP e l'IP.
I recettori muscarinici del muscolo cardiaco aumentano la permeabilità al potassio (K), iperpolarizzando così le cellule e inibendo l'attività elettrica. Si ritiene che simili meccanismi operino anche nei neuroni, dove gli analgesici oppiacei riducono l'eccitabilità attraverso l'apertura dei canali del . L'apertura del canale è prodotta dall'interazione diretta tra le subunità della proteina G e il canale, senza il coinvolgimento del secondo messaggero. Sia la subunità libera, sia il complesso delle subunità della proteina G possono essere i mediatori che controllano il canale.

Voci correlate
Collegamenti esterni
- (EN) Recettori accoppiati a proteine G, su Enciclopedia Britannica, Encyclopædia Britannica, Inc.