
Nell'essere umano e in altri mammiferi la spermatogenesi è la gametogenesi maschile, cioè il processo di maturazione delle cellule germinali maschili che avviene nei tubuli seminiferi dei testicoli. Essa ha inizio quando l'individuo ha raggiunto la pubertà, grazie allo stimolo degli ormoni FSH e testosterone.

È l'analogo dell'ovogenesi per la femmina, ma differisce da questa principalmente per le tempistiche dal momento che la produzione di spermatozoi comincia nella pubertà e dura per tutta la vita dell'individuo mentre l'ovogenesi comincia già prima della nascita salvo poi interrompersi e riprendere all'avvenuta maturazione sessuale dell'individuo per terminare alla menopausa. La spermatogenesi non è da confondere con la spermiogenesi che è la terza ed ultima fase della spermatogenesi stessa in cui avviene la differenziazione finale che porta allo sviluppo di spermatozoi maturi. Al termine della spermatogenesi solo l'80% degli spermatozoi è normale, il restante 20% è costituito da spermatozoi funzionalmente o morfologicamente anomali.
Caratteristiche generali della spermatogenesi umana
Della durata di circa 74 giorni, la spermatogenesi inizia con la divisione cellulare di cellule indifferenziate e residenti vicino alla lamina basale del tubulo seminifero (spermatogoni) che, con un susseguirsi di mitosi e meiosi, portano alla formazione di cellule mature (spermatozoi) che si distaccano dalla parte più luminale della parete tubulare. Le cellule germinali, quindi, seguono un processo che le porta dalle regioni più marginali della parete verso le regioni più apicali fino al rilascio all'interno del lume del tubulo. Gli spermatozoi, infatti, si impegnano nel lume dei tubuli seminiferi e seguono poi la strada sopra indicata fino all'epididimo dove sono in grado di stanziare anche più di sette giorni acquisendo la motilità necessaria per poter raggiungere l'ovulo da fecondare.
È importante notare come la spermatogenesi non porti alla creazione di spermatozoi completamente in grado di fecondare un ovulo: questo a causa della mancata motilità. La fase finale della maturazione avviene soltanto all'interno dell'utero o delle tube uterine attraverso la capacitazione. Questa ultima e fondamentale fase di maturazione della durata di 7 ore circa permette allo spermatozoo di perdere il rivestimento di glicoproteine e proteine seminali dalla superficie dell'acrosoma e quindi di partecipare alla reazione acrosomiale per la fusione con l'ovulo.
Aspetti di anatomia macroscopica normale dei didimi
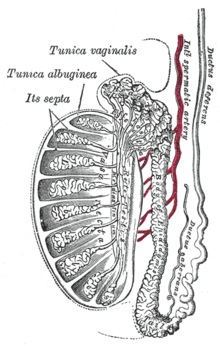

Struttura del parenchima testicolare ed epitelio germinativo
La spermatogenesi avviene nel parenchima dei testicoli (o didimi) e più precisamente nell'epitelio germinativo dei tubuli seminiferi, condotti a fondo cieco dal diametro di 150-200 µm che confluiscono nei tubuli seminiferi retti (Russell et al. 1990). I tubuli retti finiscono poi per anastomizzarsi nella rete testis da cui partono 15-20 dotti efferenti che si immettono nell'epididimo che continua poi nel canale deferente.
La parete di questi tubuli seminiferi è formata da cellule di sostegno, denominate cellule del Sertoli e svariate cellule germinali rappresentate dagli elementi che compongono le varie tappe della spermatogenesi.
Vascolarizzazione del didimo e barriera emato-testicolare
I vasi sanguigni che seguono lo stroma del testicolo (ramificazioni dell'arteria spermatica o testicolare) raggiungono l'interstizio tra i tubuli seminiferi ma mai lo spessore dei tubuli (Hinton e Turner et.al 1993). Il sangue arterioso derivante dalla circolazione è separato dall'epitelio germinativo per mezzo della barriera emato-testicolare.
Anatomia microscopica normale dei didimi
Citotipi coinvolti nella spermatogenesi

Le cellule che entrano in gioco nella spermatogenesi si dividono in due grandi gruppi: cellule germinali e cellule non germinali. Le prime sono costituite dagli spermatozoi e dai loro precursori; le seconde da cellule che non costituiscono precursori e non diventano mai gameti, ma da cellule con funzioni trofiche e regolatorie.
In una sezione trasversale di tubulo seminifero bufalo al TEM è possibile identificare nell'epitelio germinativo tre domini principali. In senso baso-apicale (cioè dalla membrana basale verso il lume del tubulo seminifero) distinguiamo tre compartimenti separati da due barriere:
- Compartimento interstiziale (con cellule mioidi e di Leydig)
- Barriera formata dalla membrana basale dell'epitelio germinativo
- Compartimento basale
2. Barriera cosiddetta "barriera emato-testicolare"
- Compartimento ad-luminale
1. Cellule germinali
Fanno parte delle cellule germinali i precursori staminali, costituiti dagli spermatogoni, spermatociti e spermatidi, e gli spermatozoi.
Spermatogoni
Gli spermatogoni sono le cellule più indifferenziate e posti verso la lamina basale del tubulo seminifero. Presentano 12 µm di diametro e sono suddivisibili in tre tipi:
-
spermatogoni A: costituiscono le cellule staminali, cioè le cellule in continua mitosi che non procedono nel processo differenziativo e si suddividono ulteriormente in:
- spermatogoni AD (da dark, scuro): sono cellule staminali o spermatogoni di riserva vista la loro posizione estremamente lontana dal lume del tubulo seminifero e dalle loro divisioni poco frequenti che possono portare o alla formazione di altri AD o alla creazione di spermatogoni AP;
- spermatogoni AP (da pale, chiaro): sono gli spermatogoni che riproducendosi portano alla formazione di spermatogoni più differenziati, fino ad arrivare agli spermatogoni B che sono l'ultima tappa prima degli spermatociti;
- spermatogoni B: sono gli spermatogoni successivi, dal punto di vista maturativo, a quelli di tipo A e costituiscono le cellule progenitrici degli spermatociti.
Dal punto di vista cromosomico, tutti gli spermatogoni sono cellule diploidi.
Spermatociti
Gli spermatociti sono le cellule che vanno incontro a meiosi e che quindi, partendo da un corredo cromosomico diploide, generano cellule aploidi. Sono due i tipi di spermatociti:
- spermatociti primari o di primo ordine: sono quelli che derivano direttamente dagli spermatogoni B e che vanno incontro alla prima divisione meiotica. Durante questa meiosi la profase dura circa 22-24 giorni ed è il motivo per cui sono le cellule maggiormente presenti nei vetrini istologici. Sono cellule diploidi.
- spermatociti secondari o di secondo ordine: sono il risultato della prima divisione meiotica degli spermatociti primari, hanno un vita molto breve e sono cellule aploidi.
Spermatidi
Gli spermatidi sono le cellule che risultano dalla seconda divisione meiotica e derivanti dagli spermatociti secondari. L'aspetto degli spermatidi è tondeggiante e non molto dissimile dagli spermatociti secondari: il nucleo è più piccolo, mentre l'apparato di Golgi presenta la vescicola acrosomiale. Gli spermatidi andranno incontro alla spermiogenesi per trasformarsi in spermatozoi.
Spermatozoi

Gli spermatozoi si generano durante la spermiogenesi a partire dagli spermatidi come spermatozoi primitivi. Essi sono costituiti da:
- una testa: occupata prevalentemente dal nucleo (costituito da cromatina altamente condensata), dall'acrosoma che ricopre circa i 2/3 del nucleo e dal quale è separato da un sottile spazio subacrosomiale e dalla fossetta d'impianto per la coda. L'acrosoma contiene una serie di molecole glicoproteiche ed enzimi proteasici fondamentali per la fecondazione.
- una coda: lunga complessivamente 55-60 µm è costituita da 4 segmenti:
- il collo che costituisce la porzione di collegamento con la testa nella fossetta d'impianto e presenta i due centrioli del sistema microtubulare;
- la parte intermedia (5 µm) dove inizia l'assonema (costituito da 9 + 2 microtubuli) che si protrarrà per tutta la coda; è avvolta da 9 fibrille esterne e una guaina formata da mitocondri avvolti a spirale;
- la parte principale (45-50 µm) costituita dall'assonema e 9 fibrille esterne circondate da una guaina fibrosa;
- la parte terminale (4-6 µm) costituita dall'assonema avvolto nella membrana plasmatica.
Sono cellule aploidi.
Cellule non germinali
Fanno parte delle cellule non germinali le cellule del Sertoli, che entrano a far parte del tubulo seminifero e, tra una cellula e l'altra, contengono le varie cellule germinali e le cellule di Leydig che hanno un ruolo estremamente marginale dal punto di vista materiale della spermatogenesi, ma fondamentale dal punto di vista regolatorio dal momento che producono il testosterone che è la principale molecola che regola l'attività delle cellule di Sertoli.
Cellule di Sertoli
Le cellule di Sertoli non fanno parte delle cellule germinali ma costituiscono le cellule di sostegno del tubulo seminifero. Esse sono infatti appoggiate sulla lamina basale del tubulo e danno direttamente nel lume. Fra una cellula e l'altra avvengono le varie tappe della spermatogenesi e, quindi, in una sezione di tubulo seminifero si potranno notare cellule di Sertoli con intercalati i vari spermatogoni, spermatociti, spermatidi e spermatozoi in via di differenziazione o maturazione.
Il ruolo delle cellule di Sertoli è puramente trofico e di supporto alla spermatogenesi, con un'importante funzione di fagocitosi nelle fasi terminali della maturazione degli spermatozoi. Le cellule di Sertoli non vanno più incontro a divisione cellulare dopo la pubertà mantenendo quindi un rapporto costante con le cellule germinali adiacenti. Nonostante la loro mancata riproduzione, sono cellule con un'altissima resistenza alle avversità (come raggi X, malnutrizione o infezioni) rispetto alle loro controparti germinali molto più delicate e quindi suscettibili agli insulti esterni.
Costituiscono la barriera emato-testicolare che impedisce al sistema immunitario di indurre l'eliminazione degli spermatociti. Questi in quanto aploidi verrebbero riconosciuti come cellule sbagliate con DNA danneggiato e quindi eliminate.
Cellule di Leydig
Le cellule di Leydig sono cellule presenti nello stroma che avvolge i tubuli seminiferi e, vista la loro lontananza dalle cellule germinali, non sono direttamente coinvolte, come le cellule di Sertoli, nella spermatogenesi. In realtà il loro ruolo è fondamentale per la loro capacità di produzione del testosterone sotto lo stimolo dell'ormone LH, una gonadotropina ipofisaria.
Il testosterone prodotto diffonde tramite l'interstizio fino alla lamina basale del tubulo dove viene assorbito dalle cellule di Sertoli, le quali sono sensibili a tale ormone, regolando così la spermatogenesi.
Fasi

La spermatogenesi si costituisce essenzialmente di 3 fasi principali: fase moltiplicativa, meiotica e differenziativa.
Nella fase moltiplicativa i precursori diploidi si replicano per mitosi producendo una parte di cellule che continueranno la moltiplicazione e una parte che procede nella fase meiotica dove si ha il passaggio da cellule diploidi a cellule aploidi che si differenzieranno in spermatozoi durante la fase finale.
Fase moltiplicativa (delle mitosi)
Nella fase moltiplicativa il principale citotipo coinvolto è lo spermatogonio, che risiede al di sopra della membrana basale che separa l'epitelio germinativo dai tessuti sottostanti. Nel corso degli ultimi decenni sono stati proposti diversi modelli di moltiplicazione e differenziazione ("commitment") delle staminali maschili umani. Secondo Bani et.Al: gli spermatogoni AD si dividono continuamente in spermatogoni AD o AP. Questi ultimi procedono differenziandosi in spermatogoni B che si evolvono in spermatociti di primo ordine, ultime cellule di questa fase.
Fase meiotica
Nella seconda fase gli spermatociti di primo ordine vanno incontro a meiosi. Durante la fase di preleptotene si ha una duplicazione di DNA che rende queste cellule già diploidi di fatto tetraploidi. Durante la prima divisione meiotica si assiste ad una profase particolarmente lunga (circa 22-24 giorni) costituita da 5 stadi:
- leptotene: con i cromosomi costituiti da un mazzetto di filamenti sottili;
- zigotene: durante il quale avviene l'appaiamento dei cromosomi omologhi e sessuali ;
- pachitene: con l'addensazione dei cromosomi sessuali che forma la cosiddetta vescicola sessuale con conseguente crossing over ;
- diplotene: in cui ha inizio la separazione dei cromosomi omologhi uniti in tetradi;
- diacinesi: dove si ha il distacco definitivo dei cromosomi omologhi.
Terminata la profase I, si ha la conclusione della prima divisione meiotica passando attraverso le sue fasi classiche: metafase I (con disposizione dei cromosomi omologhi lungo l'equatore), anafase I (con migrazione dei cromosomi ai poli cellulari) e telofase I (ricostruzione dei nuclei delle due cellule figlie). Tale divisione porta alla formazione degli spermatociti secondari che, essendo il risultato della segregazione dei cromosomi omologhi sono costituiti da un corredo aploide formato da una doppia copia di DNA (ogni cromosoma è formato da due cromatidi fratelli).
La fase meiotica si conclude con la seconda divisione meiotica che parte dagli spermatociti secondari per andare a formare gli spermatidi rotondi, cellule aploidi che non andranno più incontro a divisioni cellulari.
Fase differenziativa
La terza e ultima fase (detta anche spermiogenesi) è il momento nel quale gli spermatidi, in luogo della loro cessata divisione cellulare, cominciano a differenziarsi e a trasformarsi nei gameti attraversando tre fasi: la fase di Golgi, la fase acrosomiale e la fase di maturazione. Quest'ultima fase dura 12-24 giorni. In questo momento gli spermatidi sono cellule contenenti un nucleo, diversi mitocondri, un apparato di Golgi vicino al nucleo, un paio di centrioli, ribosomi liberi e reticolo endoplasmatico liscio. Essi nel complesso vanno incontro a fenomeni di condensazione nucleare, sviluppo del flagello, formazione dell'acrosoma e perdita di citoplasma.
Fase di Golgi
In questa prima fase nelle vescicole costituenti l'apparato di Golgi si ha un accumulo di granuli proacrosomiali che fondendosi generano un unico grande granulo acrosomiale all'interno della vescicola acrosomiale. Tutto questo avviene nel polo cellulare opposto rispetto a dove migrano i centrioli che cominciano a dare forma all'assonema di cui sarà costituito il flagello.
Fase acrosomiale
La formazione dell'acrosoma si completa con l'ampliamento della vescicola e del granulo acrosomiale che vanno ora a coprire circa la metà superiore del nucleo, il quale si sta allungando e condensando. I mitocondri si avvolgono a spirale intorno alla regione della coda chiamata parte intermedia che è la zona che durante i movimenti flagellari consumerà più energia e che quindi avrà più bisogno di mitocondri. In questa seconda fase avviene anche l'allineamento dello spermatozoo con la testa rivolta verso la parete del tubulo e il flagello in allungamento verso il lume.
Fase di maturazione
In quest'ultima fase si assiste essenzialmente alla conclusione della formazione di flagello e testa, ma soprattutto alla perdita di citoplasma, che avviene con il distacco di vescicolette, i corpi residui, che sono fagocitati dalle cellule del Sertoli. Infine si ha la spermiazione, consistente nel distacco degli spermatozoi dalle cellule di Sertoli, (e quindi dal tubulo seminifero) e il loro conseguente rilascio nel lume del tubulo.
Ciclo spermatogenico
Una caratteristica dei mammiferi maschi è la produzione continua di sperma, e pertanto la disponibilità relativamente illimitata di gameti già maturi ogni giorno.
Regolazione
Ormoni
Le cellule principali coinvolte nella regolazione della spermatogenesi sono le cellule di Sertoli, le cellule di sostegno del tubulo seminifero che entrano in contatto con tutti i vari tipi di cellule germinali. Sulle cellule di Sertoli agiscono due importanti ormoni: l'ormone follicolo-stimolante (FSH) prodotto dall'ipofisi e il testosterone prodotto dalle cellule di Leydig (le quali a loro volta producono tale ormone sotto lo stimolo dell'ormone luteinizzante (LH) ipofisario).
L'FSH, inoltre, non solo agisce direttamente sulle cellule di Sertoli per stimolarle, ma induce la produzione della proteina androgenolegante che si combina con il testosterone e lo trasporta nei tubuli seminiferi facilitando il contatto con le cellule di Sertoli stesse.
Temperatura
Fondamentale per la regolazione della spermatogenesi è anche la temperatura la quale deve essere rigorosamente di 35 °C e non di 36/37 °C come il resto dell'organismo. Il plesso pampiniforme, il plesso venoso che circonda le arterie testicolari generando uno scambio di temperatura controcorrente, la sudorazione scrotale e la contrazione dei muscoli cremasterici (con conseguente avvicinamento dei testicoli all'addome) sono alcune delle forme con cui si controlla e modifica la temperatura testicolare.
Patologie e disfunzioni

Chiaramente una qualsiasi disfunzione o blocco della spermatogenesi porta a sterilità o difficoltà nel concepimento.
Temperatura
Una delle principali problematiche che possono portare a disfunzionalità è il criptorchidismo, cioè la condizione di non completa discesa dei testicoli. Se i testicoli, che nel momento dello sviluppo prenatale (o al massimo entro i tre mesi di vita) non sono scesi dalla cavità addominale allo scroto, rimangono in una sede con una temperatura superiore a quella che permette di far avvenire la spermatogenesi (37 °C al posto di 35 °C) con una conseguente sterilità che può essere risolta con un intervento chirurgico che riporti in sede scrotale i testicoli.
Altri fattori
Tra altri fattori che possono portare a malfunzionamenti o blocchi della spermatogenesi si annoverano: radiazioni ionizzanti, farmaci, l'alcolismo, sali di cadmio, stress psichici e carenze vitaminiche (A ed E soprattutto).
La sindrome delle ciglia immobili, che può colpire anche l'apparato respiratorio, è caratterizzata da una mancanza delle proteine che permettono il movimento flagellare con conseguente immobilizzazione degli spermatozoi.
Bibliografia
- Anastasi et al., Trattato di anatomia umana, volume II, Milano, Edi.Ermes, 2007, ISBN 978-88-7051-286-1.
- Bani et al., Istologia umana, Napoli, Idelson-Gnocchi, 2012, ISBN 978-88-7947-541-9.
- Luis C. Junqueira e José Carneiro, Compendio di istologia, 5ª ed., Padova, Piccin, 2006, ISBN 88-299-1817-2.
- Keith Moore e T. V. N. Persaud, Lo sviluppo prenatale dell'uomo, Napoli, EdiSES, 2009, ISBN 978-88-7959-348-9.
Altri progetti
-
 Wikimedia Commons contiene immagini o altri file su spermatogenesi
Wikimedia Commons contiene immagini o altri file su spermatogenesi
Collegamenti esterni
- (EN) Spermatogenesi, su Enciclopedia Britannica, Encyclopædia Britannica, Inc.
| Controllo di autorità | Thesaurus BNCF 47150 · LCCN (EN) sh85126541 · BNF (FR) cb122237643 (data) · J9U (EN, HE) 987007565838605171 |
|---|