
| Briofite | |
|---|---|
 | |
| Classificazione scientifica | |
| Dominio | Eukaryota |
| Regno | Plantae |
| Divisione |
Bryophyta Schimp. |
| Classi | |
Le Bryophyta (Schimp.) sono un gruppo di embriofite prive di tessuto vascolare. Si tratta del gruppo più diversificato tra le piante non vascolari, con circa 24.000 specie. Sono conosciute comunemente come muschi.
Le briofite sono generalmente caratterizzate da tessuti vascolari non lignificati: l'assorbimento ed il trasporto dell'acqua e dei soluti necessari avviene generalmente per capillarità e interessa tutta la superficie della pianta. In alcune specie sono presenti dei cordoni di cellule con funzione conduttrice: queste strutture non sono molto efficaci e sono prive di funzione di sostegno. La mancanza di un sistema di conduzione impedisce lo sviluppo in altezza: le briofite sono piante di dimensioni piuttosto ridotte e con crescita generalmente orizzontale.
I particolari adattamenti sviluppati dalle briofite sia a livello vegetativo che a livello riproduttivo sono di grande interesse per comprendere lo studio dell'evoluzione delle piante terrestri. Le briofite, che sembrano derivare dalle alghe verdi, sono suddivise in tre gruppi: Anthocerotopsida (o antoceri), Hepatopsida (o epatiche) e Bryopsida (o muschi). Studi molecolari hanno dimostrato che le briofite sono un gruppo monofiletico.

Anthoceros agrestis

Marchantia antheridiophores


Sphagnum palustre
Caratteristiche

Differiscono dalle piante vascolari perché mancano di strutture vascolari completamente differenziate e lignificate. L'assorbimento e il trasporto dell'acqua e dei soluti avvengono soprattutto per capillarità e interessano tutta la superficie della pianta. In alcune specie sono presenti delle strutture (idroidi e leptoidi) analoghe ai tessuti vascolari xilema e floema ma comunque mai lignificati (quindi privi di funzione di sostegno) e meno efficaci nel trasporto idrico.
Il ciclo ontogenetico è aplodiplonte con prevalenza gametofitica. Lo sporofito non è autonomo ma vive a spese del gametofito fotosintetizzante. Da una spora unicellulare aploide (meiospora) si sviluppa il protonema, con dimensioni diverse a seconda delle specie. Sul protonema, aploide, si formano delle gemme da cui si sviluppano i gametofiti che a maturità portano i gametangi (archegoni e anteridi) che contengono rispettivamente i gameti femminili e maschili. Grazie alla presenza di acqua i gameti maschili raggiungono il gamete femminile rimasto all'interno dell'archegonio. Qui avviene la gamia con conseguente sviluppo dell'embrione diploide. Dall'embrione, contenuto e nutrito nell'archegonio (che fa parte del gametofito) si sviluppa lo sporofito adulto di norma non autonomo. Sullo sporofito, all'interno dello sporangio, avviene la meiosi, con dimezzamento del numero cromosomico, e si originano le meiospore.

Le Briofite presentano anche un inizio della cutinizzazione per limitare la traspirazione (stomi).
Sono importanti per la colonizzazione di nuovi ambienti, molto resistenti (reviviscenti). Le forme più conosciute sono i muschi.
La mancanza di tessuti di trasporto limita le dimensioni delle briofite: l'altezza è inferiore nella maggior parte delle specie a 6 centimetri. La più grande briofita conosciuta è un muschio dell'Australia e della Nuova Zelanda: Dawsonia superba. Questa specie raggiunge i 75 centimetri di altezza. Le più piccole briofite hanno invece dimensioni inferiori al millimetro.
Ecologia e distribuzione
Requisito fondamentale per la crescita delle briofite è quindi la presenza di acqua o comunque di condizioni di umidità per lunghi periodi. Per la crescita le briofite necessitano inoltre di un substrato di crescita relativamente stabile, di un'adeguata esposizione al sole e di un range di temperature favorevole. Molte briofite riescono a sopravvivere a periodi di siccità o congelamento attraverso un meccanismo non ancora chiaramente compreso.
Il substrato di crescita preferito è rappresentato dal terreno, dove spesso le briofite crescono sui resti di loro precedenti generazioni o di altre piante; altri substrati comuni sono le rocce nude e gli ambienti acquatici. Alcune briofite crescono su resti di animali in decomposizione, sulla superficie delle foglie, nelle saline, alla base di ciottoli di quarzo o in ambienti ricchi di rame. Altre, come le specie del genere Funaria, sono in grado di crescere su terreni bruciati in seguito ad incendi.
I modelli di distribuzione delle briofite sono simili a quelle delle piante vascolari. Alcune specie sono cosmopolite, molte crescono in zone limitate. Le briofite sono distribuite in tutto il mondo: uniche eccezioni le regioni aride, dove però possono crescere in microambienti umidi, e gli ambienti con acqua salata. Alcune briofite vivono nella zona intertidale. Le briofite sono più abbondanti nelle regioni con clima costantemente umido regioni tropicali e subtropicali; nelle regioni artiche o alpine si possono formare tappeti molto estesi che influenzano la tipologia di vegetazione e le sue dinamiche.
Riproduzione sessuata e ciclo vitale

La fase dominante del ciclo vitale delle briofite è rappresentata dal gametofito. Questo è in grado di svolgere la fotosintesi clorofilliana ed è quindi indipendente da un punto di vista nutrizionale. Lo sporofito può invece essere fotosintetico o meno: in ogni caso è sempre dipendente dal gametofito e rimane stabilmente attaccato ad esso.
Lo sporofito produce per meiosi delle spore unicellulari aploidi (meiospore). Queste spore possono restare quiescenti per lungo tempo prima in attesa delle condizioni ottimali per la germinazione. Al momento della germinazione dalle spore si sviluppa una struttura filamentosa e scarsamente differenziata detta protonema. Successivamente dal protonema si sviluppa il gametofito adulto. Sul gametofito sono presenti delle strutture piliformi, uni- o pluricellulari, dette rizoidi. La funzione dei rizoidi è quella di ancorare il gametofilo al substrato. In molte specie i gametofiti sono micorrizzati.
Sul gametofito si differenziano gli organi sessuali o gametangi. Il gametangio femminile è detto archegonio mentre quello maschile è chiamato anteridio. Esistono briofite omotalliche o monoiche e briofite eterotalliche o dioiche. Nelle prime i gametofiti portano archegoni e anteridi sullo stesso individuo; nelle seconde esistono individui che portano l'archegonio ed altre che portano gli anteridi.
Nell'archegonio è contenuta una sola cellula uovo. L'archegonio ha la forma di un fiasco, con un collo allungato ed una base rigonfia detta ventre: è in questa parte che è conservata la cellula uovo.
Nell'anteridio sono prodotti un grande numero di gameti maschili dotati di due flagelli. Alla maturità, i gameti maschili sono liberati e, sfruttando un velo d'acqua, raggiungono l'archegonio, attratti da stimoli di natura chimica. La necessità della presenza di un velo d'acqua per consentire la sopravvivenza del gamete maschile fuori dell'anteridio e durante il suo percorso fino all'archegonio è uno dei fattori che limitano la diffusione delle briofite ad ambienti umidi. Una volta giunti all'archegonio, i gameti maschili penetrano al suo interno attraverso il collo: il loro ingresso determina la produzione di una sostanza gelatinosa. Questa sostanza permette l'arrivo del gamete maschile nel ventre, dove ha luogo la fecondazione.
Lo zigote risultante dalla fecondazione ed il successivo embrione crescono nell'archegonio. Dall'embrione si sviluppa quindi lo sporofito adulto. Questo è generalmente formato da un piede, da una seta e da una capsula.
Il piede è una struttura inserita nel ventre dell'archegonio: la sua funzione è quella di permettere il passaggio delle sostanze nutrienti dal gametofito allo sporofito. La seta è un peduncolo non ramificato che consente lo sviluppo in altezza dello sporofito. La capsula è costituita essenzialmente dallo sporangio: al suo interno si formano spore aploidi. Nei muschi la capsula può contenere qualche milione di spore. A maturità la capsula si apre con meccanismi a controllo generalmente igroscopico rilasciando le spore da cui avrà inizio la successiva generazione.
Di alcune specie di briofite non si conosce lo sporofito: si tratta probabilmente di specie che hanno perso la capacità di riprodursi per via sessuata.
Limiti
Nelle briofite ogni gametofito è aploide: ogni anno forma quindi gameti con corredo genetico sempre identico. Nelle briofite omotalliche è molto probabile inoltre che i gameti maschili fecondino archegoni del medesimo gametofito generando sporofiti omozigotici. Queste condizioni determinano una grande limitazione nella possibilità di insorgenza di nuovi caratteri: in questi casi l'unica possibilità di comparsa di nuovi caratteri è legata a mutazioni spontanee.
Riproduzione asessuata
Le briofite possono riprodursi anche per via asessuata (o agamica) per frammentazione del tallo o attraverso le gemme. Queste ultime sono contenute, in alcune specie, in apposite strutture a forma di piccola scodella disposte sulla superficie del gametofito.
Le testimonianze fossili
Le testimonianze fossili riconducibili alle Briofite sono molto povere. Una delle difficoltà maggiori nel riconoscimento delle briofite nei fossili è la mancata conservazione degli sporofiti e la somiglianza dei gametofiti ad alcune piante vascolari. La prima testimonianza fossile riconducibile a briofite è in rocce provenienti dall'Argentina e datate alla prima parte dell'Ordoviciano (488-444 Ma). Si tratta di elementi simili a spore riconducibili alle epatiche. Queste testimonianze sono considerate da alcuni studiosi come la più antica prova di presenza di piante terrestri. Le prime testimonianze di muschi risalgono al Permiano (299-251 Ma). La maggior parte dei fossili di briofite è riconducibile all'Eocene (55,8-33,9 Ma). Si tratta di piante conservate in ambra con strutture molto simili a quelle attuali, tanto da permettere il riconoscimento di molte delle specie conservate. Analogo discorso per le briofite fossili risalenti al Paleogene e all'Neogene: anche in questo caso è possibile il riconoscimento della maggior parte delle specie. Le linee evolutive interne alle briofite sono quindi nate e stabilizzate in epoca molto antica.
Classificazione
Sistematica tradizionale
La classificazione tradizionale include il gruppo delle briofite nella divisione Bryophyta, che viene suddivisa in tre classi: Bryopsida, Marcanthiopsida ed Anthocerotopsida.
- I muschi (Bryopsida), con oltre 10.000 specie, sono il gruppo di briofite più diffuso. Hanno tallo cormoide con fusticino e foglioline spesso pluristratificate.
- Eubrydae: Capsula con anulus, presenza di lignina nel peristoma (nell'opercoli), specie molto diffuse.
- Andreaeobrydae: Seta di origine gametofitica
- Sphagnobrydae: Gli sfagni, praticamente immortali (la parte inferiore si degrada per dare poi nei secoli torba e carbonato-fossile); il protonema è laminare, non c'è il peristoma.
- Le epatiche (Marchantiopsida), comprendono circa 7.000 specie, a distribuzione cosmopolita con predilezione per gli ambienti umidi. Mancano del protonema, hanno rizoidi unicellulari, capsula più semplice senza columella, tra le meiospore sviluppo degli elateri per favorire la dispersione delle spore.
- Le antocerote (Anthocerotopsida) comprendono un centinaio di specie. Hanno tallo rugoso gametifitico, formato da sottili lamine, gametofito con cloroplasto e stomi, con piede che fora lo sporofito, non completamente autonomo. Da cui si pensa abbiano avuto origine le felci con Rhyniophyte estinte nel Siluriano. Quindi che siano l'anello di congiunzione con le piante vascolari superiori, perché si pensa all'ipotesi che il suo sporofito abbia potuto vivere indipendentemente dal gamentofito e che sia poi avvenuta l'inversione del ciclo aplodiplonte a fovore dello sporofito sul gametofito, come nella maggior parte delle piante vascolari superiori a partire dalle appunto dalle felci.
Filogenesi e classificazione moderna
Recenti studi sulle embriofite hanno dimostrato che il gruppo delle briofite non è monofiletico, ma parafiletico, poiché include il progenitore comune e solo alcuni dei gruppi derivati da esso. Le Polisporangiate, da cui discendono tutte le piante vascolari sono infatti escluse. Ciò ha portato all'elevazione di ciascuna delle singole classi al rango di divisione:
- Bryophyta, contenente muschi e alghe
- Marchantiophyta, a cui appartengono le epatiche
- Anthocerotophyta, con le antocerote
Esistono due probabili ipotesi sulla storia evolutiva delle piante terrestri:
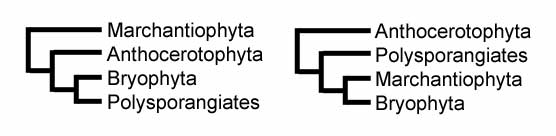
Secondo la prima ipotesi, le epatiche furono le prime a divergere, seguite dalle antocerote, mentre i muschi sono relativamente più vicine al gruppo delle polisporangiate. Nel secondo schema sono invece le antocerote a divergere per prima, seguite dalle piante vascolari, mentre muschi ed epatiche sono filogeneticamente più vicini.
Divisione Bryophyta (muschi)
- Classe Hepatopsida (epatiche)
- Ordine Calobryales
- Ordine Metzgeriales
- Ordine Jungermanniales
- Ordine Sphaerocarpales
- Ordine Monocleales
- Ordine Marchantiales
- Classe Anthocerotopsida (o antoceri)
- Ordine Anthocerotales
- Classe Bryopsida (muschi)
- Sottoclasse Andreaeidae
- Sottoclasse Sphagnidae
- Sottoclasse Tetraphidae
- Sottoclasse Polytrichidae
- Sottoclasse Buxbaumiidae
- Sottoclasse Bryidae
- Sottoclasse Archidiidae
Le briofite in Italia
Secondo le più recenti check-list la flora italiana comprende 1130 briofite. La particolare ricchezza di questa flora è evidente se si considera che in Europa sono segnalate 1690 briofite. La particolare ricchezza della flora briologica italiana è dovuta alla sua posizione geografica ed alla variabilità dei suoi ambienti.
Esiste (o esisteva) il reliquato di una specie fossile, presente sino all'ultima glaciazione: la Paludella squarrosa, nella palude di Santa Caterina di Valfurva. Sino ai primi anni ottanta essa era presente su una superficie di qualche metro quadrato, in mezzo agli sfagni. Da quegli anni non se ne ha più notizia, ma l'edilizia turistica che è avanzata occupando la palude può aver ormai cancellato definitivamente la sua esistenza.
Bibliografia
- AA. VV., 2003: Special Issue: Bryophyte. Fremontia 31 (3): 1-44. Consultabile all'indirizzo: https://web.archive.org/web/20061224155818/http://www.uam.es/informacion/asociaciones/SEB/divulgacion/Fremontia_Vol31-No3.pdf (verificato il 2 dicembre 2011).
- Glime J. M., 2006: Bryophyte Ecology. Volume 1. Physiological Ecology. Sponsored by Michigan Technological University; Botanical Society of America, International Association of Bryologists. Published on-line: http://www.bryoecol.mtu.edu/ (verificato il 2 dicembre 2011).
- Purves W. H., Sadawa D., Orians G. H. & H. C. Heller, 2001: Biologia. L'evoluzione della diversità. Zanichelli, Bologna. ISBN 88-08-09664-5.
- Konrat M. von, Shaw A. J. & K. S. Renzaglia (eds), 2010: Bryophytes: The closest living relatives of early land plants. Phytotaxa 9: 1-278. Consultabile all'indirizzo: http://www.mapress.com/phytotaxa/content/2010/pt00009.htm (Verificato il 4 dicembre 2011).
Voci correlate
Altri progetti
-
 Wikimedia Commons contiene immagini o altri file su briofita
Wikimedia Commons contiene immagini o altri file su briofita
Collegamenti esterni
- (EN) Bryophyta, su Enciclopedia Britannica, Encyclopædia Britannica, Inc.
- (EN) Bryophyta, su Fossilworks.org.
- Repertorio della flora italiana protetta, su minambiente.it (archiviato dall'url originale il 9 luglio 2011).
| Controllo di autorità | Thesaurus BNCF 4230 · LCCN (EN) sh85017380 · GND (DE) 4040211-3 · BNF (FR) cb11964814z (data) · J9U (EN, HE) 987007292551605171 · NDL (EN, JA) 00566261 |
|---|